


RSS-Feed der Bay. Landesanstalt für Wald und Forstwirtschaft abonnieren
So verpassen Sie keine Neuigkeiten mehr. Unser RSS-Feed "Nachrichten der Bayerischen Landesanstalt für Wald und Forstwirtschaft" informiert Sie kostenlos über unsere aktuellen Beiträge.
Thomas Kudernatsch, Markus Blaschke und Markus Bernhardt-Römermann
Walddynamik in der »Echinger Lohe« – LWF aktuell 122
Forschungsprojekt nimmt Vegetationsveränderungen im Naturwaldreservat unter die Lupe
Das Naturwaldreservat Echinger Lohe ist ein Relikt der einst auf der Münchener Schotterebene weit verbreiteten Lohwälder. Die Seltenheit und sicher auch die ästhetische Schönheit dieser historischen Waldform führten dazu, dass die isolierte Waldinsel schon sehr früh unter Schutz gestellt wurde. Seither wurden zahlreiche Dauerbeobachtungsflächen eingerichtet und mehrfach aufgenommen. Aufgrund dieser langen Forschungstradition ist die Echinger Lohe geradezu prädestiniert, Einflüsse von sich ändernden Umweltbedingungen auf die Vegetationszusammensetzung und Biodiversität zu untersuchen.
Ältere Studien (z. B. Seibert 1962) ordnen die Echinger Lohe den primären Eichen- Hainbuchenwäldern (Galio-Carpinetum) zu. Neuere Untersuchungen (Bernhardt- Römermann et al. 2006) deuten aber darauf hin, dass das Galio-Carpinetum in der Echinger Lohe in typischer Struktur und Artenzusammensetzung vorrangig auf Bewirtschaftungseinflüsse durch den Menschen während der letzten Jahrhunderte zurückzuführen ist und somit einen sekundären Eichen-Hainbuchenwald darstellt.
Gerberlohe, Brennholz und Waldweide
 Zoombild vorhanden
Zoombild vorhanden
Abb. 1: Frühjahrsaspekt in der Echinger Lohe. (Foto: T. Kudernatsch)
Selbst während der Weltkriege kam es noch zu Streunutzungen und somit zu einem Nährstoffaustrag und zu einer Verarmung der Böden (Kollmannsberger 1989). Erst nach der Ausweisung als Naturschutzgebiet im Jahre 1942 (seit 1978 auch Naturwaldreservat) und der damit verbundenen Einstellung der Bewirtschaftung hat der direkte anthropogene Einfluss durch nährstoffentziehende Nutzungen aufgehört.
Allerdings gibt es seit Beginn der 1960er Jahre einen konträren indirekten Einfluss: eine Vervielfachung des Angebotes an reaktivem Stickstoff durch massive Einträge aus der umgebenden Landwirtschaft sowie aus Straßenverkehr und Industrie. Seit einigen Jahren wirkt sich auch das Eschentriebsterben zunehmend auf die Waldentwicklung in der Echinger Lohe aus.
Vom Wert echter Dauerbeobachtungsflächen
Im Naturwaldreservat Echinger Lohe wurden in der Vergangenheit zahlreiche Vegetationserfassungen durchgeführt, welche für Wiederholungserhebungen geeignet sind. So wurden beispielsweise im Jahr 2003 41 über die gesamte Echinger Lohe verteilte pflanzensoziologische Dauerbeobachtungsflächen angelegt und vegetationskundlich erfasst (Bernhardt 2005). Im Rahmen des Projekts ST 330 wurden diese Flächen im Jahr 2017 erneut aufgesucht und erhoben, wobei exakt dieselbe Methodik wie bei der vorausgegangenen Erhebung angewandt worden ist, um eine Vergleichbarkeit der Ergebnisse zu gewährleisten. Durch einen Vergleich der aktuellen Aufnahmen mit der vorausgegangenen Erhebung konnten somit die im Vergleichszeitraum abgelaufenen Vegetationsveränderungen aufgezeigt und interpretiert werden.
Auch die wiederholt (1978, 2004, 2012) durchgeführten waldkundlichen Erhebungen auf der Repräsentationsfläche des Naturwaldreservats sind dazu geeignet, Vegetationsveränderungen – insbesondere im Hinblick auf die Baumartenzusammensetzung und die Bestandesstruktur – aufzuzeigen. Durch eine erneute Datenerfassung in 2017 konnte vor allem auch geprüft werden, inwieweit sich das Eschentriebsterben bereits auf die Waldentwicklung in der Echinger Lohe auswirkt.
Vegetationswandel auf allen Etagen
Baumschicht
 Zoombild vorhanden
Zoombild vorhanden
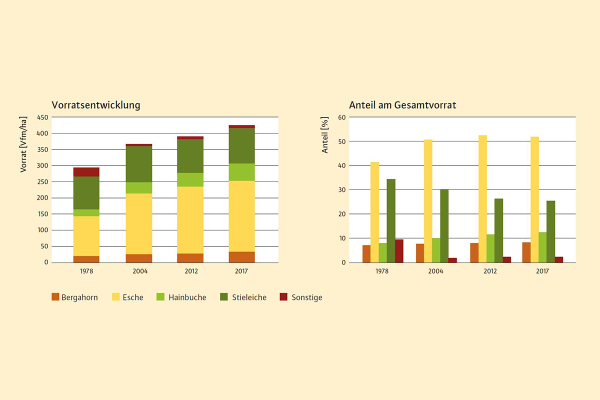
Abb. 2: Entwicklung des Vorrats auf der Repräsentationsfläche über die Zeit (1978–2017) sowie Anteile der Baumarten am Gesamtvorrat (Grafik: LWF)
Entgegen den Erwartungen hat das Eschentriebsterben bislang noch zu keinem messbaren Rückgang der Grundfläche bzw. des Vorrats der Eschen geführt. Vielmehr ist die Esche die Baumart, die – neben Hainbuche und Bergahorn – am stärksten zum beobachteten Anstieg der Grundfläche bzw. des Volumens beigetragen hat (Abbildung 2).
Dies wird sich allerdings in den nächsten Jahren vermutlich drastisch ändern, wenn große Teile der Eschen durch das Eschentriebsterben und die damit einhergehenden Folgeschädigungen absterben werden. So sind zwischenzeitlich fast alle Alt-Eschen infiziert und weisen dadurch eine stark reduzierte Belaubung sowie einen hohen Anteil an Kronentotholz auf; vereinzelte Individuen – bevorzugt innerhalb der mittleren Stärkeklassen – sind bereits gänzlich abgestorben.
Strauchschicht
Manche Arten konnten auf den Aufnahmeflächen innerhalb der Strauchschicht überhaupt nicht mehr dokumentiert werden (z. B. Stieleiche, Pfaffenhütchen, Hartriegel, Liguster, Schwarzer Holunder). Die einzigen Arten mit deutlich zunehmender Deckung innerhalb der Strauchschicht sind Hasel sowie Hainbuche.
Krautschicht
 Zoombild vorhanden
Zoombild vorhanden
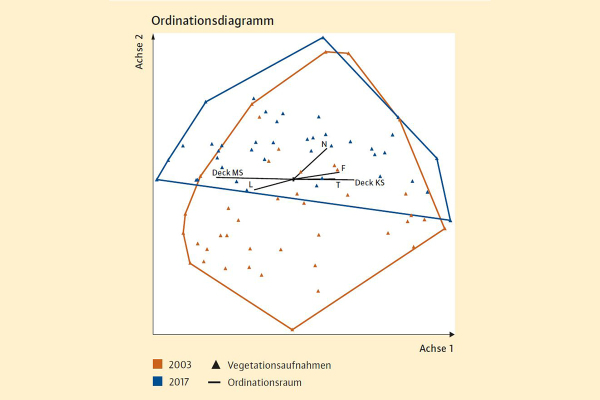
Abb. 3: Ordinationsdiagramm des Gesamtdatensatzes (82 Aufnahmen), bestehend aus jeweils 41 pflanzensozio-logischen Aufnahmen von 2003 bzw. 2017. (Grafik: LWF)
Bei der Korrespondenzanalyse werden anhand der vorkommenden Arten und deren Deckung die Positionen der Aufnahmen (bzw. der Arten) im mehrdimensionalen Raum berechnet und in einem zweidimensionalem Diagramm dargestellt. Dabei weisen Aufnahmen mit ähnlichem Artinventar eine benachbarte Positionierung auf, während floristisch unähnliche Aufnahmen weit voneinander entfernt liegen.
Vergleicht man den durch die ersten zwei Achsen beschriebenen »Ordinationsraum« der historischen Aufnahmen (2003) mit dem Ordinationsraum der aktuellen Aufnahmen (2017), erkennt man, dass sich dieser entlang der zweiten Achse von unten nach oben verschoben hat. Die floristischen Veränderungen im Vergleichszeitraum sind also überwiegend gerichtet und scheinen zu einem gewissen Grad mit einer erhöhten Stickstoff-/Nährstoffverfügbarkeit in Zusammenhang zu stehen (vgl. Stickstoff (N)-Vektor).
 Zoombild vorhanden
Zoombild vorhanden
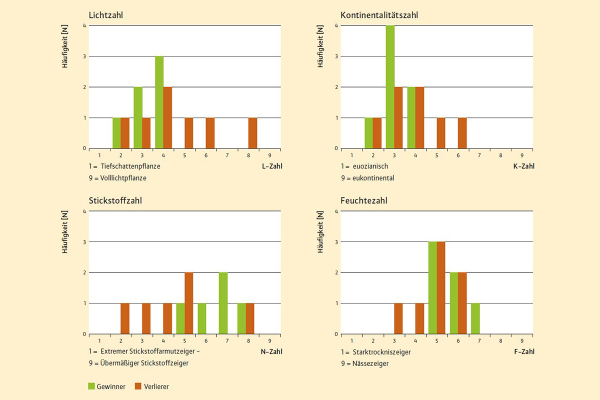
Abb. 4: Zeigerwertspektren der Arten mit deutlicher Häufigkeits-/ Deckungszunahme seit 2003. (Grafik: LWF)
So zeichnen sich die »Gewinner « durch im Schnitt niedrigere Lichtzahlen aus. Unterschiede zwischen den Arten mit positiver und denen mit negativer Reaktion gibt es auch hinsichtlich der Stickstoffzahlen. So sind die »Gewinner« zumeist Arten, die ihren Verbreitungsschwerpunkt auf meso- bis eutrophen (also gut mit Stickstoff und Nährstoffen versorgten) Standorten haben, während Arten oligotropher (magerer) Standorte eher zu den »Verlierern« zählen. Ferner sind die Arten mit Häufigkeitszunahme durch – verglichen mit den Verlierern – höhere Feuchtezahlen sowie niedrigere Kontinentalitätszahlen charakterisiert (Abbildung 4).
Was steckt dahinter? Weniger Licht
 Zoombild vorhanden
Zoombild vorhanden
Abb. 5: Die Wiesen-Schlüsselblume ist eine charakteristische Pflanzenart in lichten und wärmebegünstigten Eichen-Hainbuchenwäldern. (Foto: M. Blaschke)
Da es sich bei den Baumarten mit positiver Reaktion überwiegend um eher schattentolerante Baumarten mit großen Blättern und/oder vergleichsweise dichten Kronen (insb. Bergahorn und Hainbuche) handelt, ist von einem insgesamt geringeren Lichtangebot am Waldboden auszugehen. Darüber hinaus haben vermutlich auch die insgesamt verbesserten Wachstumsbedingungen (s. u.) zu einem gesteigerten Wachstum der Bäume und somit zu einem zunehmenden Schluss der Baumkronen beigetragen (vgl. auch Bernhardt-Römermann et al. 2009).
Dass es – insbesondere in Waldbeständen mit abnehmender Bewirtschaftungsintensität – über die Zeit zu einem dichteren Kronenschluss und damit einer Zunahme schattenverträglicher Arten (bzw. einer Abnahme lichtbedürftiger Arten) in der Krautschicht kommt, konnten auch andere Studien innerhalb Europa zeigen (z. B. Verheyen et al. 2012; Heinrichs & Schmidt 2017). Auch der beobachtete Rückgang von Arten mit höheren Kontinentalitätszahlen dürfte mit einem dichteren Kronenschluss in ursächlichem Zusammenhang stehen. Da die Kontinentalitätszahl vor allem das Ausmaß von Temperaturschwankungen abbildet, könnte der Rückgang auf ein insgesamt ausgeglicheneres Bestandesklima hindeuten, wodurch Ein- und Ausstrahlungseffekte verringert werden.
Mehr Stickstoff
Einen wesentlichen Faktor stellen sicherlich anthropogene Stickstoffeinträge dar, die zu etwa 40 % aus der Verbrennung (Straßenverkehr, Industrie und Hausbrand) und zu etwa 60 % aus landwirtschaftlichen Quellen stammen (BMUB 2017). So sind die Stickstoffeinträge in die Waldökosysteme seit Jahrzehnten zu hoch, eine zunehmende Stickstoffsättigung der Wälder ist bereits vielerorts zu beobachten (Dietrich et al. 2018). Da die Echinger Lohe als isolierte Waldinsel von intensiv landwirtschaftlich genutzten Flächen umschlossen ist und auch die nahe gelegenen, in Hauptwindrichtung vorgelagerten Autobahnen einen bedeutsamen Stickstoff-Emittenten darstellen, ist ein Einfluss von Stickstoffeinträgen auf die Vegetation sehr wahrscheinlich.
Neben anthropogenen Stickstoffeinträgen kann die Zunahme von Arten mit höheren Stickstoffzahlen aber auch auf die Einstellung der einst intensiven, teilweise nährstoffentziehenden Bewirtschaftung zurückgeführt werden (s. o.). Die nach Ausbleiben dieser intensiven Nutzungen einsetzende Regeneration der Böden (z. B. Humusakkumulation) und die damit verbundene bessere Stickstoffverfügbarkeit (aber auch Wasserverfügbarkeit; vgl. höhere Feuchtezahlen der Gewinner) stellt somit eine weitere wahrscheinliche Ursache für die floristischen Veränderungen innerhalb der Echinger Lohe dar.
Die Verbesserung des Stickstoffangebots für die Pflanzen könnte zusätzlich noch durch den Bestockungswandel (s. o.) beschleunigt worden sein, da sich die Baumarten mit positiver Reaktion (wie z. B. Esche und Bergahorn) durch eine gut zersetzbare Streu auszeichnen (Verheyen et al. 2012). Dies ist ein gutes Beispiel dafür, dass ein Standortfaktor (wie hier die Stickstoffverfügbarkeit) im Verlauf der Zeit durch verschiedene Wirkfaktoren beeinflusst und verändert werden kann (Jantsch et al. 2013).
Wildverbiss
 Zoombild vorhanden
Zoombild vorhanden
Abb. 6: Im NWR Echinger Lohe sind die negativen Auswirkungen des Wildverbisses nicht zu übersehen. (Foto: T. Kudernatsch)
So beschreibt schon Albrecht (1989), dass die hohen Rehwildbestände aus der umliegenden landwirtschaftlichen Flur ihre bevorzugten Einstandsgebiete in der Echinger Lohe haben und dadurch die Bodenvegetation sowie die nächste Generation der Waldbäume (und Sträucher) stark beeinträchtigen. Durch diesen intensiven Wildeinfluss sowie das geringe Lichtangebot ist es offenbar vielen Gehölz-Individuen nicht mehr möglich, von der Krautschicht in die Strauchschicht einzuwachsen (Abbildung 6).
Zusammenfassung
Als wesentliche primäre Wirkfaktoren konnten insbesondere die Einstellung der Bewirtschaftung des einst intensiv genutzten Lohwaldes, anthropogene Stickstoffeinträge sowie ein intensiver Wildeinfluss identifiziert werden. Diese Faktoren führen mittel- bis langfristig zu einem Verlust der charakteristischen Arten- und Strukturvielfalt des sekundären Labkraut-Eichen- Hainbuchenwaldes und somit zu einem Wandel der Biodiversität. Prognostiziert man die ablaufenden Prozesse in die Zukunft, ist davon auszugehen, dass sich Artenzusammensetzung, Struktur und Diversität des Schutzgutes Echinger Lohe noch deutlicher als bisher verändern werden.
Dieses Beispiel zeigt einmal mehr, dass ein Erhalt der kulturhistorisch entstandenen, durch Mangel und Störung konditionierten Biodiversität sekundärer Eichenwälder nur möglich ist, wenn gezielte Managementmaßnahmen ergriffen werden.
Projekt
Literatur
- Albrecht, L. (1989): Die Echinger Lohe – ein Naturwaldreservat. In: Gemeinde Eching (Hrsg.): Garchinger Heide und Echinger Lohe
- Bernhardt, M. (2005): Reaktionen der Waldbodenvegetation auf erhöhte Stickstoffeinträge: Analyse und Vorhersage von Vegetationsveränderungen anhand von funktionellen Merkmalen. Dissertation TU München
- Bernhardt-Römermann, M.; Östreicher, S.; Fischer, A.; Kudernatsch, T.; Pfadenhauer, J. (2006): Das Galio-Carpinetum im Münchener Raum – Ergebnis früherer Bewirtschaftung? Tuexenia 26: S. 27–36
- Bernhardt-Römermann, M.; Kirchner, M.; Kudernatsch, T.; Jakobi, G.; Fischer, A. (2006a): Changed vegetation composition in coniferous forests near to motorways in Southern Germany: The effects of traffic-born pollution. Environmental Pollution 143: S. 572–581
- Bernhardt-Römermann, M.; Pfadenhauer, J.; Östreicher, S.; Fischer, A. (2009): Stickstoffbedingte Vegetationsveränderungen in einem Eichen-Hainbuchenwald – Ergebnisse aus 18 Jahren Dauerbeobachtung. Forstarchiv 80: S. 181–188
- BMUB – Bundesministerium für Umwelt, Naturschutz, Bau und Reaktorsicherheit (Hrsg.) (2017): Stickstoffeintrag in die Biosphäre; erster Stickstoffbericht der Bundesregierung. Berlin
- Dietrich, H.-P.; Raspe, S.; Zimmermann, L.; Wauer, A.; Köhler, D.; Schubert, A.; Stiegler, J.; Blum, U.; Kudernatsch, T.; Klemmt, H.- J. (2018): Umwelt- und Standortsbedingungen in raschem Wandel. LWF aktuell 117: S. 6–10
- Ellenberg, H.; Weber, H. E.; Düll, R.; Wirth, V.; Werner, W. (2001): Zeigerwerte von Pflanzen in Mitteleuropa. 3. Aufl. Scr. Geobot. 18: S. 1–262
- Ewald, J.; Hennekens, S.; Conrad, S.; Wohlgemuth, T.; Jansen, F.; Jenssen, M.; Cornelis, J.; Michiels, H.; Kayser, J.; Chytrý, M.; Gégout, J.; Breuer, M.; Abs, C.; Walentowski, H.; Starlinger, F.; Godefroid, S. (2013): Spatial and temporal patterns of Ellenberg nutrient values in forests of Germany and adjacent regions - a survey based on phytosociological databases. Tuexenia 33: S. 93–109
- Fischer, A. (1999): Sukzessionsforschung: Stand und Entwicklung. Ber. Reinh.-Tüxen-Ges. 11: S. 157–177
- Heinrichs, S.; Schmidt, W. (2017): Biotic homogenization of herb layer composition between two contrasting beech forest communities on limestone over 50 years. Applied Vegetation Science 20: S. 271–281
- Hill, M. O.; Gauch, H. G. (1980): Detrended correspondence analysis: an improved ordination technique. Vegetatio 42: S. 47–58
- Jantsch, M. C.; Fischer, A.; Fischer, H. S.; Winter, S. (2013): Shift in Plant Species Composition Reveals Environmental Changes During the Last Decades: A Long-Term Study in Beech (Fagus sylvativa) Forests in Bavaria, Germany. Folia Geobot. 48/4: S. 467–491
- Kollmannsberger, F. (1989): Die Echinger Lohe. – In: Gemeinde Eching (Hrsg.): Garchinger Heide und Echinger Lohe
- Seibert, P. (1962): Die Auenvegetation an der Isar nördlich von München und ihre Beeinflussung durch den Menschen. Landschaftspflege und Vegetationskunde 3: S. 1–123
- Verheyen, K.; Baeten, L.; De Frenne, P.; Bernhardt-Römermann, M.; Brunet, J.; Cornelis, J.; Decocq, G.; Dierschke, H.; Eriksson, O.; Hédl, R.; Heinken, T.; Hermy, M.; Hommel, P.W.F.M.; Kirby, K.; Naaf, T.; Peterken, G.; Petřík, P.; Pfadenhauer, J.; Van Calster, H.; Verstraeten, G. (2012): Driving factors behind the eutrophication signal in understorey plant communities of deciduous temperate forests. Journal of Ecology. 100: S. 352–365